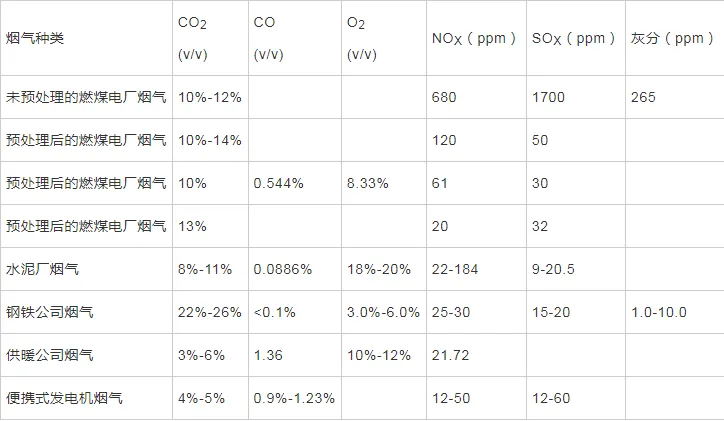
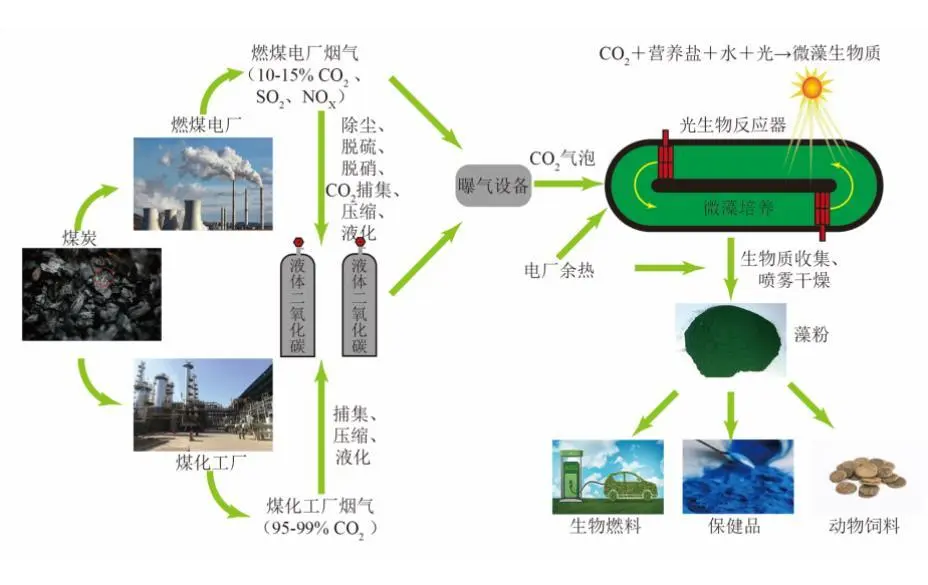
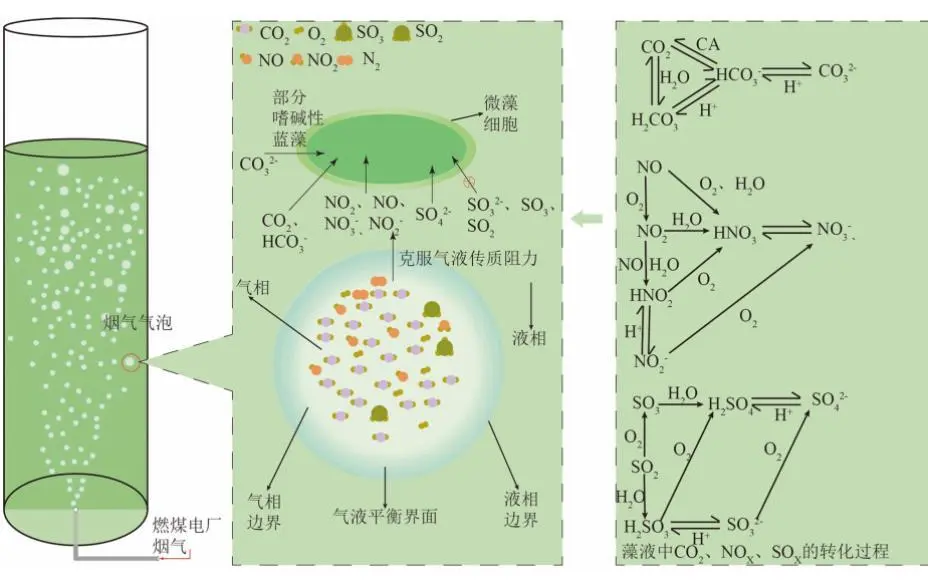
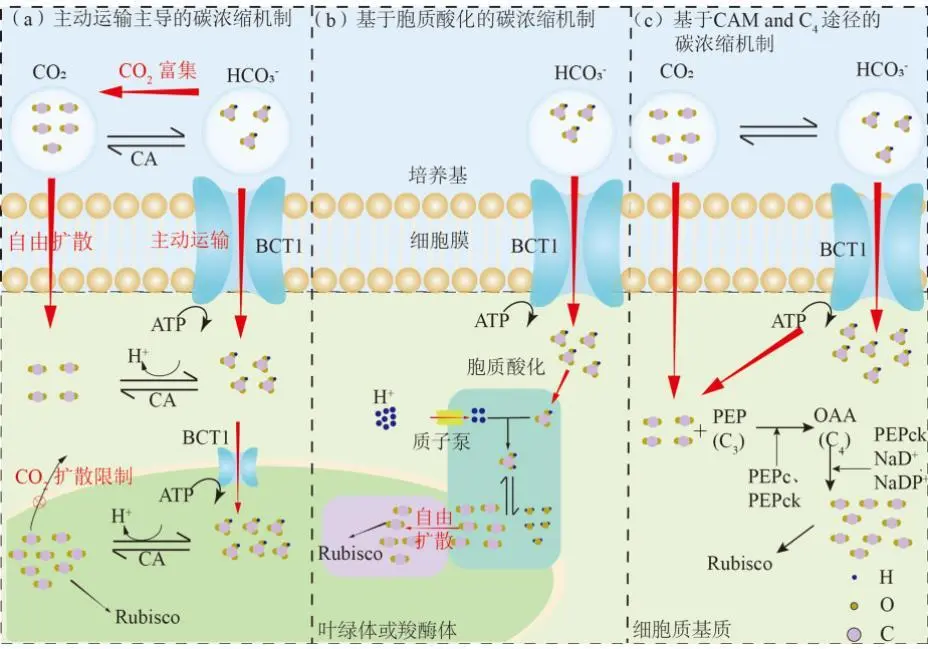
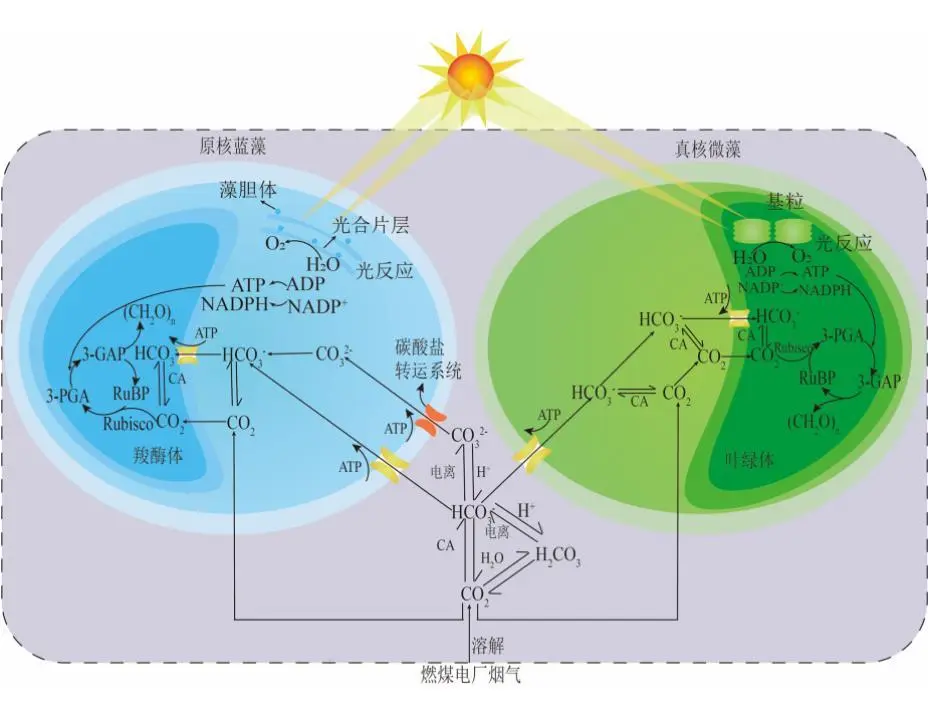
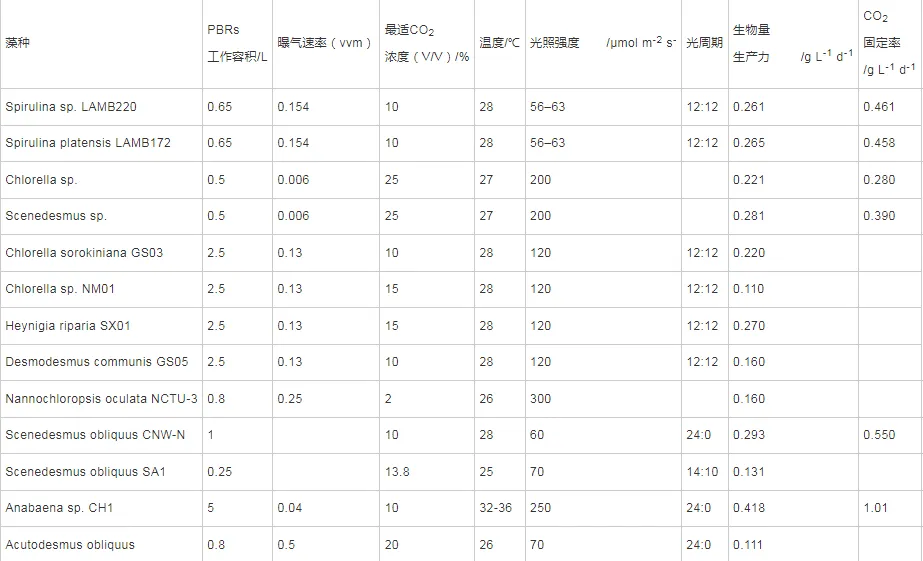
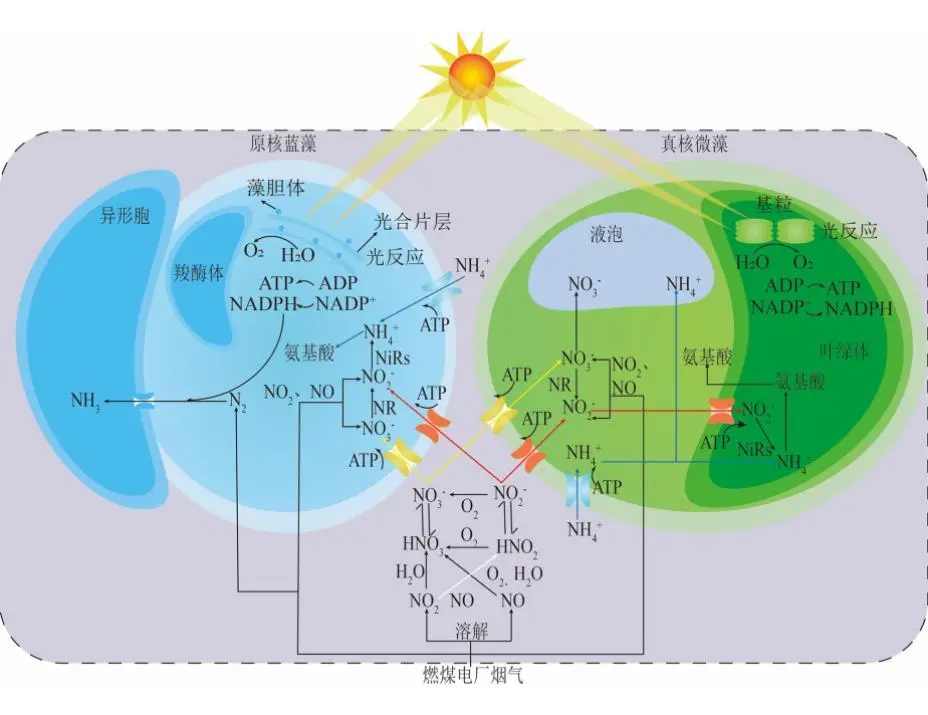
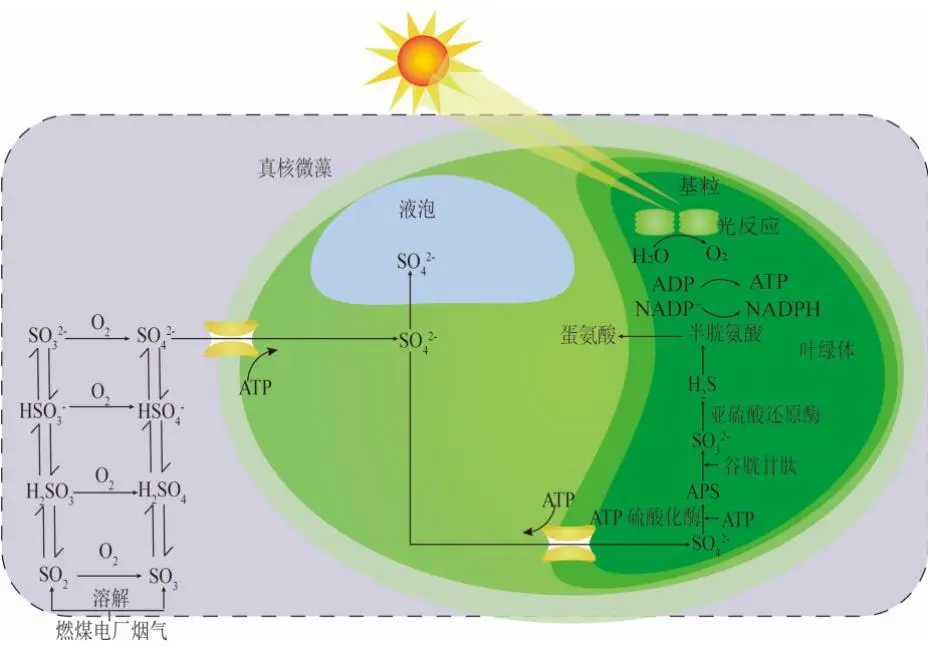
2022年7月,空气中CO2浓度已经达到417.44 ppm ,这远远高于工业化前280 ppm的水平;据预测,按现有的CO2排放速度,到21世纪末空气中CO2浓度将达到950 ppm。国际能源署在2023年发布的《CO2 Emissions in 2022》中指出, 2022年全球碳排放量达到了368亿吨的新高。作为世界上最大的碳排放国,我国的碳排放总量从2001年加入世界贸易组织时的032亿吨攀升至2022的114.77亿吨,预测2026年将提前达到峰值131亿吨。2020年,国家主席习近平在第七十五届联合国大会.上提出了我国碳达峰、碳中和的宏伟战略,积极应对大气CO2浓度上升造成的全球气候变化。 为阻止大气中CO2浓度持续攀升和降低大气中CO2浓度,可采取以下两个策略:一是限制CO2的产生。但是,对于发展中国家,限制CO2产生可能会制约其社会经济的发展。二是通过碳捕捉、封存及利用技术,减少CO2的排放。碳捕捉与封存最早由Nordhaus和Marchetti提出,旨在实现长期的CO2地质封存或海洋封存。根据全球碳捕捉与封存研究院发布的《Global Status of CCS Report ( 2023)》, 全球目前共有41个正在运营的CCS大型设施、26个在建设施,这些CCS设施每年可以贡献3.61亿吨的CO2固定量,约为全球碳总排放量的1%。为了降低CO2捕捉、压缩、储存、运输产生的成本,早期研究者们将捕捉获得的CO2用于提高石油和天然气的采收率,在实现CO2封存的同时对其加以利用,进而降低碳捕获成本。这种策略即碳捕捉、利用及封存( Carbon capture, uiliation and storage, CCUS)。当前,除地质利用之外, CO2的利用方式还包含物理利用(制备干冰等)、化学利用(加氢合成甲醇等)、生物利用(微藻固碳等)。与CCUS不同 ,碳捕捉及利用( Carbon capture and utiliation, CCU )技术中, CO2不会被长期储存,而是被转化为再生燃料以替代化石燃料或驱动其他工业过程,如作为碳源用于培养微藻。然而,在这些利用CO2的工业过程中,能源消耗会排放大量CO2 ,甚至超过该过程的CO2捕获量。因此,该技术并不具备大幅度减少CO2排放的潜力,更侧重于构建一种循环经济。 就从大气中去除CO2而言, CCS是最合适的选择,但CCS设施的运行需要相应的经济支持,可持续性较低。从人类社会经济发展的角度, CCUS在CCS和CCU间找到了一个恰当的平衡点,是最具可持续发展的减碳策略。 在各种CO2利用方式中,微藻固碳具有极高的潜力。微藻生长速率快,对CO2的生物固定能力是陆生植物的50倍,而且每固定1.83 kg CO2就可以产出1 kg微藻生物质。CO2经微藻固定后可以转化为细胞内的有机碳骨架,生成脂类、蛋白质和碳水化合物等有机物,可用于开发燃料、动物饲料、生物塑料等产品,实现CO2的高值化利用。此外,微藻不仅可以在固定CO2的同时回收废水中的氨、磷元素,还不参与耕地的竞争111。本课题组通过优化培养条件使得小球藻 MBFJNU-1对原始沼液总氮、离子型氨氮和总磷的去除效率分别达到了74%、78%和87% ,并且在通入500 mL min-1 5% CO2的条件下获得了454 mgL-1 d-1的CO2固定速率。在此基础上,在装液量3000 L (液面高度30 cm )的跑道池中探究了在中试规模下小球藻MBFJNU-1对原始猪沼液的氮磷去除效率以及CO2固定能力,在通入2L min-1 3% CO2的条件下,获得了478.5 mgL-1的生物量、63.2 mg L1d-1的CO2固定率、82.1%的总氮去除率、28 4%的总磷去除率以及37.1%的化学需氧量( COD )去除率;所获得的小球藻生物质在15 L反应釜中经酯交换获得了83%的生物柴油转化率,为微藻碳减排协同废水处理提供了强有力的依据。可观,微藻固碳技术是实现双碳”目标极具潜力的策略。微藻固碳的发展可以分为两个阶段。1991年 2008年,研究者们揭示了微藻的碳浓缩机制( CCM) ,证明了微藻固碳的理论可行性。2009年后,微藻固碳的研究重点转向其产业化应用,旨在通过研究微藻高值化固碳的机理,胁迫微藻富集高附加值产物;或通过协同优化光和碳的供应,提高微藻的CO2固定效率和光合效率,进而提高微藻固碳规模化应用的经济可行性。本文从烟气组分、光照、温度、pH等因 素对微藻生长和固碳的影响以及提高微藻固碳能力的策略等方面介绍了近年来国内外微藻固定燃煤电厂烟气CO2的研究进展。 1.1烟气的组分及其含量 燃煤电厂发电过程中,煤炭在空气中燃烧产生的烟气中CO2浓度通常不高于15% , N2浓度不低于70%。煤炭中硫元素几乎全部都会被氧化成SO2 ,仅有少部分会进一步氧化成SO3,排放这些硫氧化物( SOx )会形成硫酸型酸雨,腐蚀黑色金属和植被;燃煤电厂烟气的氨氧化( NOx )中NO约占90%~95%、NO2约占5%~10% ,煤炭中的微量含氮有机物燃烧以及氮气在电厂锅炉的高温和催化剂条件下和氧气饭应均会生成NO ,其中5%的NO会和O2进一步作用生成NO2 ,或发生一系列复杂反应生成微量的N2O3、N2O4、N2O及N2O5,往大气中排放NOx会形成硝酸型酸雨和光化学烟雾。如表1所示,不同的工业烟气中CO2浓度存在较大差异,通过预处理可以有效降低燃煤电厂烟气中SOx、NOx的含量。我国环保部、国家发改委和国家能源局在2015年联合印发的《全面实施燃煤电厂超低排放和节能改造工作方案》中指出, 全国所有具备改造条件的燃煤电厂力争在2020年实现污染物超低排放,即电厂尾气中SO2、NOx、烟尘排放浓度分别不超过35mgm-3、50 mg m-3、10 mg m-31。目前,该方案已经圆满达成,全国各地燃煤电厂烟气污染物含量均低于排放阈值。 表1 不同烟气的组分差异 1.2燃煤电厂烟气CO2的捕集工艺 随着“双碳”战略的提出,在完善燃煤电厂烟气脱硫脱硝工艺的同时,烟气的脱碳也势在必行。燃煤电厂的碳捕获技术可以分为燃烧前碳捕集、富氧燃烧以及燃烧后碳捕集,。燃烧前捕集以氧气或水蒸气等为气化剂,在高温下通过化学反应( C+H2O→CO+H2 )将煤转化为CO、H2的混合物, CO和H2O经催化剂作用在高温高压下进一步反应(CO+H2O→CO2+H2)生成CO2 ( 20% 40% )和H2 ( 40% -60% ),最终通 过分离获得高浓度的CO2和H2 , H2可以用于火力发电也可以用于其他行业。富氧燃烧则是将煤在纯氧中燃烧,由于没有N2的存在,这种方式产生的烟气中CO2的浓度可达80% -98%,而且几乎不产生氮氧化物。燃烧后捕集是指采用化学或物理方法对电厂烟气中的CO2进行选择性富集,由于电厂烟气中CO2浓度相对较低( <15% ), CO2分离难度相对富氧燃烧和燃烧前捕集较大。但是,该技术相对成熟、简单,不需要制氧设备,只需要对现有烟气系统进行小幅改造就可以达到脱碳要求。因此,燃烧后捕集技术是最具潜力的碳捕集技术。CO2的捕集方法一般有化学吸收法、 物理吸收法、吸附分离法、膜分离法以及低温液化分离法等。目前,单乙醇胺等胺基碱性溶剂介导的化学吸收法在燃烧后捕集中最具应用前景,也是第一个商业化的碳捕集技术。在物理吸收法中,溶剂和CO2不通过化学键连接, CO2的溶解度取决于其分压,因此,该方法更适合吸收富养燃烧产生的高浓度CO2或在高压条件下分离CO2和H2。吸附分离法采用碳基或高分子材料、二氧化硅、沸石、金属有机框架( MOFs )等固体吸附剂从烟气中捕获CO2;因其操作成本低、能耗少,被认为有望能替代胺基碱性溶剂吸收法。但是,在低CO2压力下,吸附剂对CO2的吸附容量和选择性较低,吸附法的适用性仍然有限。膜分离法用于分离烟气中的CO2和N2是可行的,但该方法需要高压环境,更适用 于燃烧前碳捕集。低温液化分离法是捕获CO2的绿色技术,但是,维持液化所需的低温条件( <-70℃ )所需的能源成本限制了该方法的应用。无论何种方法,捕获的CO2都需要经纯化( >95% )、压缩、储存后加以利用。 如图1 ,电烟气或被捕集的CO2由曝气器通入微藻培养体系,在光照条件下,微藻将CO2转化为其自身生物质,完成CO2的封存。在一个培养周期后,光性物反应器( PBRs )中部分微藻被收集、干燥成藻粉,最后加工为高附加值产物,实现CO2的高值化利用; PBRs中剩余的微藻在补充氨源、磷源、钾盐和痕量金元素后重新开始快速生长,实现微藻对CO2的连续固定。 图1 微藻减排燃煤电厂烟气CO2工艺示意图 由于CO2和H20分子极性不同, CO2在水中的溶解容易饱和;在一个大气压的条件下,100%CO2在纯水中只能产生3964 mg L1的可溶性无机碳( DIC ) 。CO2在藻液中的溶解饱和后,气泡中的CO2会大量逸散到大气中,造成微藻的固碳效率大幅降低。往藻液中添加醇胺类化合物、碳酸盐、氦溶液和氨基酸盐等可以有效提高藻液的CO2负载量,进而提高微藻的固碳效率。在微藻固碳系统中, CO2的捕集效率受到工艺和环境的影响。有研究表明,碱性添加剂介导的CO2吸收微藻固定法对CO2的捕集效率可达70%-80%。 1.3烟气CO2对微藻生长和固碳性能的影响 1.3.1 CO2在微藻悬浮液中的传质 如图2 ,在鼓泡通气的过程中, CO2需要通过气相薄膜和液相薄膜才能溶解到藻液中,被微藻利用。CO2经式(1)至式(4)的反应形成CO2(aq)、H2CO3、CO32-、HCO3等形式的DIC ;在这些DIC中, HCO3和CO2 (aq )是微藻的理想碳源,也有报道称部分嗜碱性蓝藻可以吸收C032-。 图2 微藻悬浮液中CO2. NOx和SOx的传质过程 如图3(a) , CO2和HCO3进入微藻细胞的方式主要有3种: ( 1 ) HCO3借助膜上的主动转运蛋白BCT1 ,通过耗能的主动运输进入细胞; ( 2 )通过碳酸酐酶( CA )将HCO3转化为CO2实现局部CO2富集后扩散进入细胞; ( 3 )利用浓度差为推动力以CO2的形式直接扩散穿过细胞膜。如图4 ,进入细胞后, CO2会被运输到叶绿体或者羧酶体中的核酮糖- 1,5-二磷酸羧化酶( Rubisco )的活性位点处,通过卡尔文循环被微藻同化。 1.3.2碳浓缩机制( CCM ) Rubisco是微藻光合作用的关键酶。该酶具有羧化酶的催化特性,即催化暗反应中1,5-二磷酸核糖( RuBP )与CO2反应生成3-磷酸甘油酸( 3-PGA )的过程。Rubisco也具有加氧酶的催化特性,即催化RuBP与O2反应形成PGA和磷酸乙醇酸;该催化过程发生了光呼吸,减少了光合产物的形成与积累,造成了光能的损失。Rubisco表现何种酶的催 化特性取决于其活性位点处CO2和O2的浓度,由于Rubisco对CO2的亲和性较低,在低浓度CO2的时候, Rubisco会催化RuBP与02反应,导致了较低的固碳速。 由于空气中CO2浓度低且CO2在水中溶解度低,微藻的天然生存环境中CO2浓度极低。为了适应低浓度CO2的生存环境,绝大多 数微藻都进化出了碳浓缩机制( CCM) , CCM本质上就是增加Rubisco活性位点附近CO2浓度的过程,富集后CO2浓度可达环境的1000倍。如图3 ,微藻的CCM分为三大类: (1)依靠HCO3单向转运蛋白BCT1主导的主动运输。这种CCM在蓝藻中尤为明显,由于基质CA的缺乏,蓝 藻细胞质基质的无机碳大都以HCO3的形式存在,难以转化为CO2,这就使得蓝藻胞内CO2浓度处于极低的水平。在胞内低浓度CO2的诱导下,该转运蛋白的HCO3-亲和性将显著增强以大量转运HCO3- ,进而实现碳浓缩; (2) Rubisco周围胞质溶胶酸化推动的CO2富集。Rubisco仅能以CO2的形式利用无机碳 ,在CA的不参与的情况下, HCO3向CO2的转化缺少强有力的推动,胞质溶胶的酸化会产生大量H+ ,将HCO3转化为高浓度CO2后以自由扩散的形式运输至Rubisco活性位点,实现碳浓缩; (3)基于C4和景天酸代谢( CAM )途径的碳浓缩机制。这种机制中CO2首先与磷酸烯醇式丙酮酸(PEP)合成C4中间体草酰乙酸(OAA) ,而后中间体在Rubisco的活性位点处发生脱羧放出CO2 ,从而实现Rubisco处CO2的富集。 图3 微藻的碳浓缩机制 1.3.3 CO2浓度对微藻生长和固碳能力的影响 从微藻的生理特性上看, CO2是微藻光合作用关键酶Rubisco的底物。当该酶活性位点处CO2浓度较高时,该酶表现出较强的羧化酶催化特性,有利于CO2的固定和有机物的积累。从CO2运输形式来看,低浓度CO2时, CO2需要先转化为HCO3再通过主动运输进入微藻细胞;而高浓度CO2时, CO2可以利用细胞内外的CO2浓度差以自由扩散的方式进入微藻细胞。相较于低浓度CO2 ,高浓度CO2条件下, CO2分子的运输方式更为高效而且减少了相应的能量投入,利于生物质积累。但是,随着大量CO2涌入细胞质基质,胞内CA会催化CO2和H20作用产生HCO3和大量H+ ,导致基质酸化。胞内低pH会抑制PS I ( 光合系统I ) 的活性;此时, PS I (光合系统| )的循环光合磷酸化更加活跃,产生更多的ATP用于质子泵,将H运出藻细胞,维持胞内适宜的pH水平。从对CCM的影响来看已有研究发现:在高浓度CO2供应下,微藻细胞内CCM的部分基因表达水平会下调。这可能是因为此时微藻细胞可直接吸收CO2 ,而无需进行碳浓缩。值得注意的是,朱颜霞等人的研究指出,极高浓度( 99.99% )的CO2会使得CCM相关基因和HCO;主动转运蛋白基因的表达水平升高,呈现出与极低浓度CO2时相同的现象;这可能是为了防止极高浓度的CO2通过浓度差大量扩散入细胞,造成细胞损伤,微藻主动改变碳吸收的形式,导致Rubscio活性位点处CO2浓度反而不足,从而激活CCM机制,促进HCO3主动转运。从产物合成的类型来看, CO2的通入会升高微藻培养基的碳氮比,使得微藻多糖合成增加而蛋白质合成减少。在低浓度CO2时,微藻倾向于合成饱和脂肪酸,而在高浓度CO2时则倾向于积累不饱和脂肪酸,这可能是因为适宜浓度的CO2会增强酰基去饱和酶ACP和乙酰辅酶A合成酶的表达。 图4 原核蓝藻和真核微藻的CO2固定机制 适宜浓度的CO2可以促进微藻生长和固碳是毋庸置疑的,但是,通入高CO2浓度的气体培养微藻时,微藻的生长通常会受到抑制。如表2所示,在这些微藻固碳相关研究中,随着气体中CO2浓度的升高,微藻的生物量生产力和固碳速率先上升至最高值,后逐步下降,最适合微藻生长的CO2浓度一般不超过10%。 当然,不同藻种的CO2耐受能力存在明显的差异,有少数微藻能耐受高CO2浓度( 20%-50% ) 。对于高浓度CO2抑制微藻生长的机制,研究者们持不同的观点。学者人在研究中指出高CO2浓度本身不会对微藻的生长产生影响,引入高浓度CO2后导致的胞外pH下降才会抑制微藻的生长学者人的CA敲除实验也同样印证了低pH对微藻的抑制作用。不同的是, 学者人认为高浓度CO2分子对微藻产生的毒害作用应是独立于胞外pH下降造成的抑制作用之外的,往藻液中持续通入高浓度CO2后形成的超高碳环境会使得微拟球藻( Nannochloropsis oceanica CCMP1779 )细胞呈现出许多与人类肝细胞相似的特征;例如, D-氨基半乳糖这样的强氧化性分子在细胞中积累,促使脂质过氧化、漂白光合色素引发DNA损伤,最终导致细胞死亡。 表2 不同微藻耐受CO2能力的比较 值得注意的是,能够抑制微藻生长的高浓度CO2是指藻液中的CO2浓度而非气体中所具有的CO2体积比,往藻液中通入高CO2浓度的气体并不一定会在 藻液中富集高浓度CO2。例如,将CO2以10 vvm、10%CO2和0.01 vvm、100%CO2两种曝 气钫式加入藻液,10 vvm、10%CO2会在藻液中 富集更高浓度的CO2。因此,纯CO2可以直接用于培养微藻。但是,应该根据微藻培养体系的温度、pH、 光照等因素,调整合适的气体流量。此外,如式(5) , CO2在藻液中转化是一个复杂的过程,DIC存在形式取决于藻液pH ;酸性或中性条件下, DIC大以CO2(aq )形式存在。由于高浓度CO2分子的毒性可能在酸性或中性条件下才会体现,亦或是其本身造成的抑制作用被pH下降造成的影响所掩盖,高浓度CO2分子本身是否对微藻有抑制作用仍需更深入研究。 1.4 NO对微藻生长的影响 氮是微藻细胞生长和代谢所需的重要元素,约占微藻细胞干重的10%。微藻可利用的无机氮源有NO2、NO3、NH4+、N2等。燃煤电厂烟气中的NO和NO2溶于水后可以通过自由扩散进入微藻细胞,在细胞质中被转化为NO2及NO3- ,也可以在微藻培养基中发生如式(6)至式(10)的反应转化为离子型的无机氮进而被微藻吸收。NO是一种亲脂、疏水的小分子化合物,难溶于水,仅能在培养基中存在溶解氧的时候才能大量和水发生反应,通过螯合作用可以增大其在水相中的溶解度,增加被微藻去除的效率;在氧气浓度适合的条件下也可以通过发生式(11)的反应转化为更易溶于水的NO2。NO2是易溶于水的红棕色气体,不同条件下可以和水作用生成HNO3、HNO2 , NO和NO2也可以发生归中反应生成HNO2 ; HNO3、HNO2会中和微藻培养基中的碱性物质并形成NO3、NO2 ,从而降低微藻生存环境的pH。如图5 , NO3、NO2可以通过细胞膜.上的硝酸盐转运蛋白( NRT2 )运输至细胞内。在细胞质基质中,部分NO3会在硝酸还原酶( NR )的催化下还原为NO2 ; NO2则会被转运至叶绿体中并在亚硝酸还原( NiRs )的催化下进一步还原为NH4* ;然后通过一系列的生化反应将一部分NH4*同化为氨基酸、核苷酸、叶绿素等含氮有机物,余下的NO3和NH4则会被储存到液泡中15.581。 可见, NO可以作为氮源被微藻吸收;但是,高浓度N0产生的亚硝酸盐会造成细胞膜渗漏。此外, NO因结构与O2相似,也可以成为细胞色素氧化酶的错误底物而可逆地抑制细胞呼吸。目前的研究报道中,添加缓冲液用以维持pH稳定或添加外源生长调节激素是提高微藻对烟气中NOx耐受能力的有效手段,如Wang等通过添加外源的亚精胺提高了微藻对抗NO的能力。 图5 原核蓝藻和真核微藻的NOx同化机制 1.5 SOx对微藻生长的影响 硫是藻细胞内L半胱氨酸、L-蛋氨酸、谷胱甘肽等功能性代谢物的基本组成元素,约占生物量干重的0.3%-0.5%。烟气中的SOx都是易溶于水的气体,可以与水通过式(12)至式(14)的反应生成H2SO4 ,也可以通过式(15)的反应被NO2氧化为H2SO4 ,最终通过中和反应生成S042。如图6 ,微藻主要是通过硫酸盐转运蛋白将溶液中SO42输送至细胞内, -部分SO42进入叶绿体而余下SO4-则将会被储存在液泡中, S042需在叶绿体中被ATP活化为5-腺苷酸( APS )才能被进一步还原成SO3-和H2S ,最后同化为半胱氨酸、甲硫氨酸等。与NOx相似,烟气中的SOx会和水反应生成H2SO4和H2SO3 ,造成微藻培养基酸化,严重时会抑制微藻生长。SO2与培养 基作用生成的SO32可以通过竞争性抑制Rubisco ,影响光合作用;或通过阻碍线粒体ATP生成影响呼吸作用。亚硫酸盐的细胞毒性受到环境影响,酸性条件下亚硫酸盐表现出强毒性,碱性条件下则几乎没有毒性。此外,当较高浓度的亚硫酸氢盐转化为硫酸盐时会形成超氧阴离子、羟基自由基、过氧化氢等强氧化性分子,会氧化膜脂和漂白叶绿素,抑制微藻生长。 在微藻固定燃煤电厂烟气CO2过程中,烟气中SO2通常与CO2-起通入微藻培养液中,SO2气体在水中的溶解度远高于CO2。初始阶段气体中所含的SO2遇水快速溶解,与水作用生成H2SO3并电离出H+ ,使得pH降低,抑制了气泡中CO2的溶解,不利于CO2分子被微藻细胞吸收利用。研究表明,通入0.1 vvm含有15%CO2的模拟烟气培养小球藻时,添加0-300 ppm SO2会轻微抑制小球藻生长;随着SO2浓度升高, SO2的抑制作用增强,当SO2浓度 高于400 ppm时小球藻的生长被完全抑制。微藻对SO2的耐受浓度阈值也受微藻生物量浓度、通气速率等影响。我国电厂烟气污染物超低排放战略实施后,电厂烟气SO2的浓度普遍低于35 mg m-3 (约为14 ppm) , 在适宜的曝气速率下大多数微藻都可以耐受其带来的影响。 图6 真核微藻的SOx同化机制 另外,从光照对微藻固定CO2的影响(光在光生物反应器( PBRs )中的传输过程、光照强度的影响、光暗交替频率的影响、光周期的影响、光波长的影响)、温度和pH对微藻固碳的影响(ph值、温度)等层面分析,得出提高CO2固定效率和光合效率的策略,其中包括: 1选育高固碳能力的藻种 2增强CO2的传质 3优化光供给策略 三个方面。 微藻是最主要的初级生产者之一,在自然界的碳循环中起着重要的作用。微藻固碳对环境友好, 可持续性高,是CCUS中最具潜力的CO2利用方式之一。高固碳能力的藻种是高效利用CO2的先决条件,微藻对烟气CO2的耐受机理以及高效同化CO2的机制需进一步揭示 ,为微藻固碳提供理论支撑。目前,大规模应用中微藻的固碳能力和生物质产量较实验室水平还有差距。由于气泡与藻液间的高CO2传质阻力以及PBRs中不均匀的光分布,微藻对CO2和光能的利用率较低。通过设计新型曝气器、优化PBRs的结构、采用更合理的光供应策略可以有效解决这一问题。由于微藻采收、脱水干燥、提取藻油需耗费大量能源,利用微藻藻油生产生物柴油的成本高于化石燃料,竞争力弱。因此,微藻生物质的精炼方向正往更高附加值的产物转变,如藻蓝蛋白、多不饱和脂肪酸等。此外,高值化利用后产生的微藻藻渣也可以作为生物能源等产品的原料,实现微藻生物质的分层次多级利用,将CO2的价值最大化地提高。可见,微藻固碳技术在实现我国双碳'目标的过程中具有极大的应用前景。 文章来源: 仅作科普 侵删