

高效生物生产碳氢化合物是解决石油等液体燃料短缺的有效手段之一,而微藻油是生产可持续生物燃料的可靠选择。布朗葡萄藻( Botryococcus braunii)是一种由单细胞组成的不定形群体绿藻,能够积累大量的碳氢化合物,最高含量可高达其干重的75%,因而受到广泛的关注。近年来随着对葡萄藻生物学特性和生长生理的不断深入研究,提高了其规模化培养及其碳氢化合物工业化生产的可行性。
本文从生物学特性、碳氢化合物台成途径及调控因子、多组学研究和规模化培养技术几方面简单地叙述了布朗葡萄藻作为新型产油微藻生产碳氢化合物的潜力,为探索利用布朗葡萄藻大规模工业化生产生物燃料提供参考,从而加速该微藻资源的开发利用。

01 布朗葡萄藻生物学特性
布朗葡
布朗葡萄藻是一种由梨状单细胞组成的不定形群体绿藻,细胞通常长7- 10μm ,宽4-6μm,以球形花束的形式排列,广泛分布于世界各地的淡水和微咸水湖泊、水库、池塘甚至是暂时性的水洼中。布朗葡萄藻细胞被大量含碳氢化合物的胞外聚合物连接包裹在细胞外侧区域,多糖原纤维延伸形成鞘状结构。每次藻细胞分裂后,从细胞壁延伸出来的鞘原纤维会被新的原纤维所取代,而细胞外薄层的原纤维则不断累积形成鞘的其余部分而不被替换。这种方式可能是布朗葡萄藻在不断进行细胞分裂的同时,还能够保留储存在细胞基质外的液态碳氢化合物的一种独特生长策。
布朗葡萄藻因其与大部分动、植物或细菌不同,能够积累大量的长链碳氢化合物(烷烃或烯烃)的特性而受到广泛的关注。而根据其合成碳氢化合物的不同,可将布朗葡萄藻分为四个族,分别为A族、B族、L族和S族。利用分子生物学技术对布朗葡萄藻分类的现代研究表明,通过18S rRNA或18S rDNA基因序列鉴定出的主要分支与通过碳氢化合物类型进行的分类相对应。A族布朗葡萄藻主要产生奇数碳直链烯烃,碳数从C23到C33,碳氢化合物分子中有两个或三个双键;B族主要产生不饱和三萜类碳氢化合物,主要为C30 -C37的葡萄藻烯( botryococcene )和角鲨烯( squalene )及二者的甲基化衍生物;L族产生C40四萜类碳氢化合物,主要为番茄二烯( lycopadiene )及其衍生物。S族布朗葡萄藻是最新被鉴定分离出的一族,主要合成C18环氧正构烷烃和C2o饱和正构烷烃。
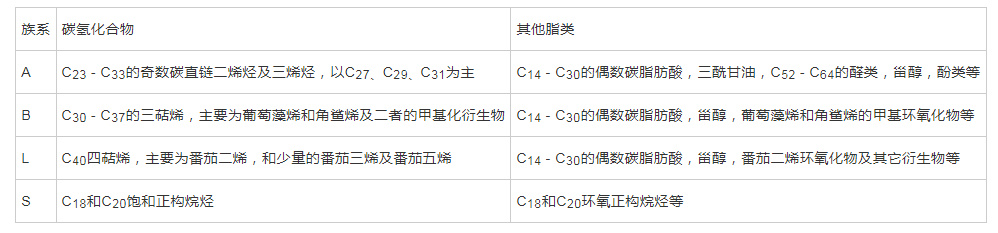 表1 已知的四族布朗葡萄藻的碳氢化合物及其他主要脂类产物
表1 已知的四族布朗葡萄藻的碳氢化合物及其他主要脂类产物
除此之外,不同族间的差异还在于其稳定生长期或受营养盐胁迫后的藻细胞颜色变化。布朗葡萄藻在大量累积油脂时,叶绿素含量降低, A族由绿色变为淡黄色;B族由绿色变为橘黄色;L族由绿色变为橙红色(图1 )。不同颜色变化的差异是由于B族和L族布朗葡萄藻的细胞中积累了酮类胡萝卜素(角黄素、棘皮酮等)。B族和L族累积的酮、萜类化合物也使得其比累积脂肪烃类化合物的A族和S 族布朗葡萄藻更耐细菌攻击。此外,不同族的布朗葡萄藻也具有不同的形态特征。例如,A族和B族藻细胞相对更大,其平均个体长度可达13 μm ,宽度可达8 μm ,而L族藻细胞长度约为9 μm ,宽度约为5 um。
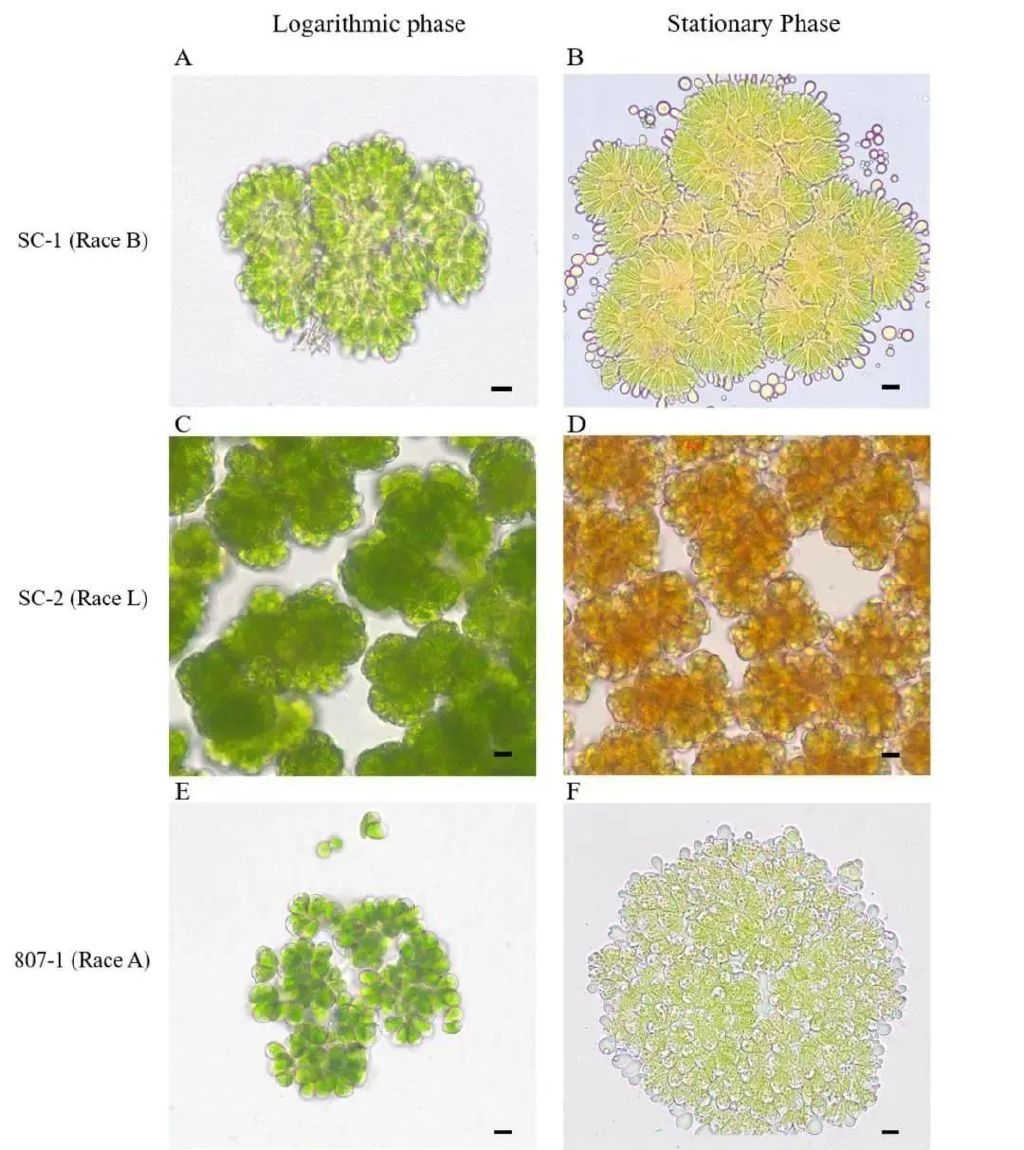 图1 三个不同族的布朗葡萄藻(807-1 (A族)来自SAG藻种库: SC-1 (B族)和SC-2 (L族)本实验室采集分离于华南地区的水域)(左列为藻细胞处于对数生长期,细胞绿色:右列为藻细胞处于平台期,图中比例尺大小为5μm)
图1 三个不同族的布朗葡萄藻(807-1 (A族)来自SAG藻种库: SC-1 (B族)和SC-2 (L族)本实验室采集分离于华南地区的水域)(左列为藻细胞处于对数生长期,细胞绿色:右列为藻细胞处于平台期,图中比例尺大小为5μm)
02 布朗葡萄藻烷烃、烯烃合成途径
几乎很少有生物能够产生高浓度的液态烃。在异养菌中,耐盐细菌弗氏弧菌(Vibrio furnissii)能产生高达其生物质干重60%的胞外烃,主要是C15H32至C24Hso的饱和烷烃。然而,该物种的潜在毒性阻碍了其在工业上的开发应用。破囊壶菌(Aurantiochytrium sp.)菌株18W-13a,能够积累达其生物质干重19.8%的角鲨烯。但是该菌株的优化培养基含有蛋白胨、酵母提取物和葡萄糖,对于角鲨烯的商业化生产来说成本非常昂贵。而布朗葡萄藻的碳氢化合物含量可达其生物量干重的61%甚至更多,且大部分是在细胞外侧基质中积累的,而不是在细胞内或培养基中积累的。这一特性使得其碳氢化合物比其他在细胞内脂质体中积累三酰甘油的微藻更易提取分离,目前被研究以提高葡萄藻碳氢化合物的生产率,并降低其工业生产的成本。
001从CO2固定到重要的有机中间代谢产物合成
微藻能够利用CO2和带电阴离子HCO3进行光合作用,但不能够利用碳酸盐离子CO32-作为碳源。由于CO2在水中的扩散率低于在空气中的扩散率,以及微藻生活的水体环境导致的CO2与水的复杂电离平衡,自然界中微藻的主要碳源通常是HCO3而非CO2。无机碳通过卡尔文循环生成葡萄糖,再经糖酵解途径生成3-磷酸甘油醛(G3P)及丙酮酸(PA),并在有氧条件下经丙酮酸脱氢酶系(PDHC)进一步氧化分解转化为乙酰辅酶A(Acyl-CoA)。在布朗葡萄藻中,所有碳氢化合物都是由上述几种初级代谢物作为前体物质合成产生的。
002奇数碳链状脂肪烃的合成
藻类的脂肪酸合成通常在叶绿体中进行。乙酰辅酶A经由乙酰辅酶A羧化酶(ACC)催化生成丙二酰辅酶A(Malonyl-CoA),丙二酰CoA是脂肪酸生物合成的通用碳前体,与ACP结合,由酰基载体蛋白转酰基酶(MCAT)、脂肪酸合酶(FAS)等一系列酶催化,在脂肪酸合成途径中每次循环便添加1分子丙二酰CoA,经多次循环可合成棕榈酰ACP(C16:0-ACP)直至硬脂酰ACP(C18:0-ACP)。随后可以通过碳链延长酶加碳延长,用于合成长链脂肪酸;或通过脂肪酸去饱和酶(FADS)和硫脂酶(FAT)催化水解,合成多不饱和脂肪酸,如棕榈油酸(C16:1)和油酸(C18:1)。
通过放射性标记示踪剂实验,发现油酸可能是布朗葡萄藻合成二烯烃和三烯烃的前体物质;有实验结果表明,脂肪醛类化合物可能是碳氢化合物合成的直接前体。研究从A族布朗葡萄藻中成功分离出了钴卟啉脱羧酶(CPE),纯化的醛脱羧酶可以催化十八醛脱羰为十七烷。进一步的研究也证实了通过脱羰途径生产脂肪烃类碳氢化合物的可能,利用NADPH作为还原辅助因子将长链的酰基CoA或酰基ACP还原成脂肪醛,由脂酰还原酶(FACR)、脱羧酶(CPE)催化脱羧基反应。该酶在豌豆中被发现可催化脱羰基反应,后被证实也存在于A族布朗葡萄藻中。
003 异戊二烯类衍生物的合成
布朗葡萄藻萜类化合物的合成过程中,首先需要在细胞内通过乙酰辅酶A合成异戊烯基焦磷酸(IPP)和二甲基烯丙基焦磷酸(DMAPP),这是萜类生物合成的前体物质,它们可以通过甲羟戊酸(mevalonate pathway,MVA)和非甲羟戊酸(non-mevalonate pathway,non-MVA)两种途径合成。过去,IPP和DMAPP这两种物质一直被认为在所有生物中均是通过MVA途径合成的,由乙酰辅酶A作为底物经一系列酶促反应先合成中间产物甲羟戊酸,再缩合生成IPP及DMAPP。但近年来对于藻类的研究发现,部分藻类缺失MVA途径的基因。它们是通过2-C-甲基-D-赤藓糖醇-4-磷酸途径(2-C-methyl-D-erythritol 4-phosphate,MEP)(通常也被称为非甲羟戊酸途径)合成IPP和DMAPP的。对B族布朗葡萄藻的碳标记实验也表明,其MEP通路基因表达活跃,三萜类化合物并非由MVA途径合成。在MEP途径中,由糖酵解产生的丙酮酸和G3P作为底物,在1-脱氧-D-木酮糖-5-磷酸合成酶(DXS)和还原异构酶(DXR)的催化下合成中间产物MEP。MEP经过一系列酶催化反应合成2-C-甲基赤藓醇-2,4-环焦磷酸(MECPP),在羟甲基丁烯基磷酸合酶(HDS)的催化下生成4-羟基-3-甲基-2-丁烯基焦磷酸(HMBPP),最后由羟甲基丁烯基磷酸还原酶(HDR)还原为IPP及DMAPP。IPP和DMAPP之间可以通过异戊二烯焦磷酸异构酶(ID1)催化而相互转化,并通过缩合和去磷酸化生成萜类化合物,不同的焦磷酸盐合成酶催化DMAPP与IPP合成牻井儿基焦磷酸盐(GPP,C10);IPP或DMAPP与GPP合成法尼基焦磷酸盐(FPP,C15);两分子的GPP或一分子的IPP和FPP缩合生成牻牛儿基拢牛儿基焦磷酸(GGPP(C20)。萜类化合物在细胞质或线粒体中单独或协同合成,每一种萜类缩合酶都对其焦磷酸盐底物具有特异性,如单/倍半/二/三/四萜烯,都有其特异的合成酶。
布朗葡萄藻合成的异戊二烯类化合物,如角鲨烯、葡萄藻烯和番茄二烯等都是通过萜类生物合成途径合成的。角鲨烯合酶(SQS)在真核生物中广泛存在,分别催化两步反应:
(1)两分子FPP偶联缩合形成一个稳定的环丙基羰基二磷酸中间体(前角鲨烯二磷酸盐,PSPP);
(2)PSPP的去磷酸化、环裂解、碳键重组异构化及NADPH还原,从而合成角鲨烯。葡萄藻烯是角鲨烯的异构体,通过与角鲨烯生物合成类似的途径合成,但在第二步反应时由不同的酶催化。角鲨烯是通过PSPP的C1'-1键还原重排产生的,而葡萄藻烯是通过C1'-3键的还原重排产生的。Okada和Devarenne根据已知的角鲨烯合酶序列,对一株B族布朗葡萄藻CDNA文库进行同源性筛选,分离出了一种葡萄藻烯合酶(BSS),但在大肠杆菌中表达的BSS仅合成角鲨烯,而不合成葡萄藻烯。角鲨烯和葡萄藻烯随后会通过三萜甲基转移酶(TMT)被甲基化,其甲基化的量是可变的,形成不同的甲基化产物。而L族布朗葡萄藻合成番茄二烯则是通过一种与角鲨烯合酶结构相似的番茄二烯合酶(LOS),其合成过程与角鲨烯合成过程类似,通过催化两分子GGPP缩合形成C40中间体,再进行还原去磷酸化等合成番茄二烯。
我们对本实验室采集、纯化分离自华南地区水域的一株L族布朗葡萄藻SC-2的碳氢化合物研究发现,其碳氢化合物主要成分为番茄二烯,还含有番茄三烯(lycopatriene,C4oH76)以及少量的番茄五烯(lycopapentaene,C4oH72),这两种成分应为番茄二烯合成途径的中间产物。
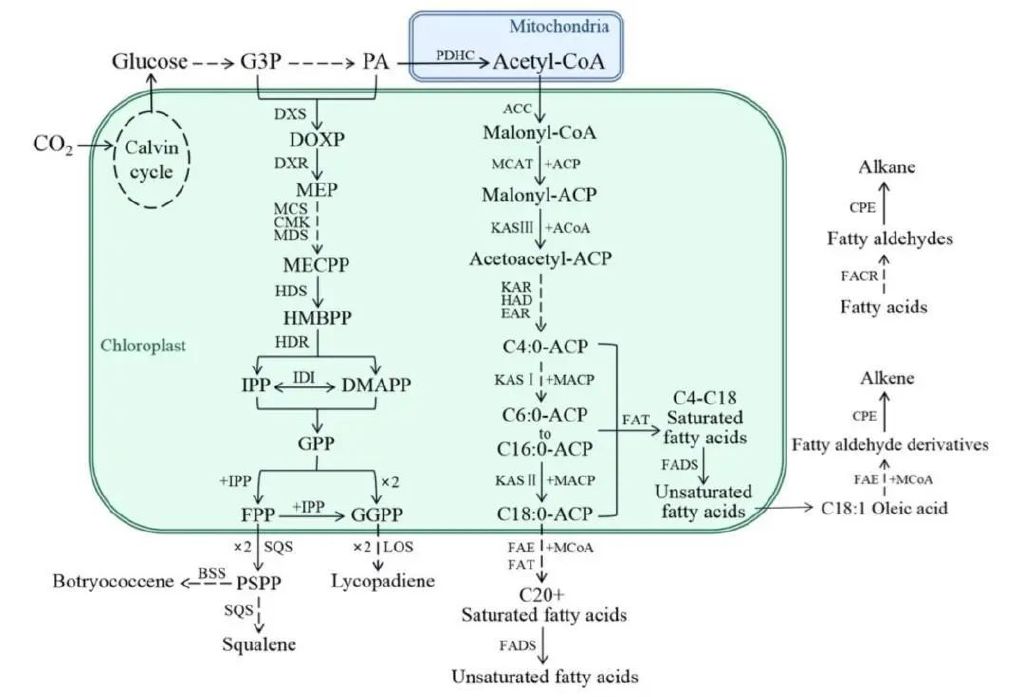 图2 布朗葡萄藻脂肪酸、烷烃和烯烃及萜类烯烃合成途径
图2 布朗葡萄藻脂肪酸、烷烃和烯烃及萜类烯烃合成途径
03 布朗葡萄的多组学研究
为了提高藻细胞代谢产物产量,传统的方法主要是通过优化培养基和培养工艺,以及改进下游提取工艺等。通过代谢工程对藻类进行改良的技术仍处于不完善的阶段,需要进一步探索。使用组学手段(基因组学、蛋白质组学、代谢组学、转录组学等)可以深入了解藻类的基本生物过程,为藻类生物质能源的发展提供了新的思路。
001基因组学的研究
基因组学分析提供了关于藻类生物合成和代谢的信息,为提高其作为细胞工厂的生产力提供了蓝图。随着基因组新测序技术的出现、成本的降低和质量的提高,已发表的藻类基因组序列的数量正在不断增加。学者用流式细胞仪分别对几株A族、B族和L族的布朗葡萄藻基因组大小进行测序,发现A族Yamanaka藻株的全基因组大小约为166.0±0.4 Mb,B族葡萄藻的基因组大小约为166.2±2.2 Mb,与A族葡萄藻的基因组大小几乎相同,然而L族葡萄藻Songkla Nakarin藻株的全基因组大小则显著高于A族和B族,约为211.3±1.7 Mb,不同族之间的差异有助于绘制布朗葡萄藻的进化关系。另外,基因组大小和基因数量之间似乎存在普遍的相关性,而L族布朗葡萄藻可能含有更多的基因。Richard等人利用古基因组测序技术对化石中的早期布朗葡萄藻进行了DNA纯化与分析,进一步了解了已灭绝的与现存的布朗葡萄藻之间的系统发育关系。比较基因组学能够检测物种内和物种间的基因和功能变异,藻类的基因组分析能够鉴定出新的生物合成基因簇,用于生产多种化合物,并应用于生物医学和工业微生物领域[60]。Zou等人对一株布朗葡萄藻的线粒体基因组进行了测序组装及注释,其大小为84,583 bp,包含31个蛋白编码基因、23个tRNA基因和3个rRNA基因。其中蛋白编码基因包括atp、cox、nad、cob、tatc及核糖体蛋白等重要基因。Olga等人也得到了一株B族葡萄藻叶绿体及线粒体的基因组序列,长度分别为156,498bp和129,356bp。其中叶绿体基因组与已发表的布朗葡萄藻SAG807-1的叶绿体基因组(172.83kb)相比略小,但具有大致相同的顺序和保守基因,包含105个推测的蛋白质编码基因,31个tRNA和3个rRNA基因,能够注释的有81个蛋白质编码基因,包括32个与光合作用相关的基因。线粒体基因组包括23个tRNA、3个rRNA和43个推测的蛋白编码基因,其中18个基因与呼吸作用相关。其包含的大量开放阅读框(orf)和丰富的非编码区使它成为目前为止已知的绿藻门中最大的线粒体基因组。叶绿体和线粒体细胞器在细胞的能量代谢中起着重要的作用,因此获得布朗葡萄藻细胞器的基因组数据也有助于进一步的遗传学研究以及分类和系统发育分析。
002 代谢组及脂质组学的研究
布朗葡萄藻含有大量较高应用价值的代谢产物,由于藻类代谢过程的复杂性,可以通过代谢组学帮助分析细胞内的代谢物及其合成途径,还可以比较不同族不同培养条件下的代谢产物。学者对布朗葡萄藻CCAP807/2进行了全面系统的代谢组分析,并通过GC-MS首次分析了培养过程中藻细胞内代谢物的波动,发现在线性生长后期可以观察到高达细胞干重63%的碳水化合物产量增加,而高达细胞干重24%的细胞外碳氢化合物积累主要发生在稳定生长期,揭示了液态烃和碳水化合物的含量和代谢阶段的差异,表明该株布朗葡萄藻更倾向于合成碳水化合物而非烃类化合物。在进行藻类共培养时对某A族葡萄藻的次级代谢产物分析发现,在培养条件改变的情况下,其合成的烃类物质也会有微小的改变,如甲基化等官能团的变化。除了代谢组外,还可以通过脂质组学对布朗葡萄藻代谢产物中的总脂质部分进行分析。这不仅有助于识别已知的脂质成分,还有助于检测新的脂质,用于更好地了解不同族间葡萄藻的脂质代谢,以优化其生物柴油的生产力[67]。对布朗葡萄藻的脂质组进行分析后,发现除了常见的存在于绿藻内的脂肪酸外,还在几种布朗葡萄藻中鉴定出超长链脂肪酸(VLCFAs);其中最丰富的体外游离脂肪酸(C28:2、C28:1和C28:0)包含于中性脂类(三酰基甘油及二酰基甘油)和磷脂(磷脂酸及磷脂酰胆碱)中;并首次分离出了含有VLCFAs的磷脂酰胆碱。进行代谢产物的分析可以更好地了解布朗葡萄藻细胞的整体生化特征,并为生产应用提供有价值的参考,所获得的数据也可能对其他组学研究至关重要,如转录组学和蛋白质组学。
003转录组学的研究
转录组分析在布朗葡萄藻的组学研究中最为常见,它是一种分子工具,可以深入了解基因表达,并获取微藻的功能细节。转录组分析提供了有关活跃编码基因组区域以及发育代谢等分子方面的信息,可用于代谢途径和网络的重构。布朗葡萄藻最为人们关注的就是其合成碳氢化合物的特性,目前有大量关于其碳氢化合物代谢途径、分子调控机制及相关酶基因的转录组研究。通过转录组分析,确定了布朗葡萄藻的萜类合成是通过MEP途径而非MVA途径。有研究人员从B族布朗葡萄藻中分离鉴定出了角鲨烯合成酶(BSS)基因,并发现其在基因组中只有一个拷贝。这表明角鲨烯和葡萄藻烯的生物合成可能存在合成酶基因的协同表达,或者可能存在一个独特的控制角鲨烯合成酶和葡萄藻烯合成酶活性的肽。Niehaus等人分离鉴定出了与BSS同源的蛋白质,角鲨烯合酶样蛋白SSL-1、SSL-2和SSL-3。SSL-1作为PSPP合成酶发挥作用,SSL-2催化合成角鲨烯,SSL-3催化合成葡萄藻烯。纯化的SSL-1在体外既不能合成葡萄藻烯也不能产生角鲨烯,相反,当SSL-1添加到布朗葡萄藻细胞提取物中时,能够刺激葡萄藻烯的产生。David和Motohide等人均对B族葡萄藻的全转录组进行了研究分析,重建了由初级代谢物合成萜类液态烃的途径,以及将光合碳转化为四萜类胡萝卜素、二萜类和叶绿素等的异戊二烯途径。有研究成功在L族葡萄藻中分离出了推测为合成番茄二烯前体的合成酶,对不同族间的转录组比较分析表明,葡萄藻烯合成酶和番茄二烯合成酶分别对布朗葡萄藻的B族和L族具有特异性。由L族葡萄藻转录组分析重构了番茄二烯的生物合成途径,发现番茄二烯合成酶(LOS)除了GGPP外,还能够利用FPP作为底物,可以单独或组合合成六种不同的C30、C35和C40链长的碳氢化合物。
为了更好地理解生物分子层面的特征,可以整合来自基因组学、转录组学、蛋白质组学和代谢组学等组学进行综合分析,有助于了解整个系统和特定控制条件(如氮缺乏胁迫)引起的生物学变化。
众所周知,与石油相比,藻类生产生物燃料的成本较高,是因为其需要培养、提取,加工成本高。进一步的改良可以使其在经济上可行,并为其提供工业规模生产的准备。组学技术和系统生物学的发展将藻类作为微生物细胞工厂应用的想法推到了前沿,有希望操纵藻类的代谢途径,以提高重要工业产品的产量。
04 总结
尽管布朗葡萄藻具有高产烃的优势,但在大规模生产应用方面仍存在一些局限性和挑战。布朗葡萄藻生物量增长缓慢,生长周期长,且易受到环境影响。微藻的生长速率、生物量产量和生化组成主要受光照、pH、CO2、温度、营养盐含量等参数的影响,规模化培养是获得生物质原料的最重要方面之一。因此,商业化大规模培养技术的选择和优化是非常重要的。
微藻规模化培养技术主要分为开放式和封闭式两种系统,开放式系统是室外设施,如池塘、跑道池和浅循环系统等,其养殖成本较低,但易受污染,藻类的产量及质量也会受到影响。跑道池和带搅拌臂的开放式池塘是目前微藻最常用的开放式培养系统,但这种培养方式适合具有特殊生长条件的藻类商业生产,如小球藻和螺旋藻,不适用于布朗葡萄藻。为了克服开放式培养系统带来的限制,可以采用封闭系统进行规模化藻类培养。封闭系统也被称为光生物反应器,包括垂直柱状反应器、气升式反应器、螺旋或环形管状反应器、平板和塑料袋等,其应用更加灵活,可通反应器设计、配置、运行可持续性、占地空间要求等因素进行优化,更容易控制光照强度、二氧化碳含量、营养盐水平、温度等参数,不易受到污染,从而提高藻类产量,降低收获成本。
 图3 常见的微湿养殖系统
图3 常见的微湿养殖系统
布朗葡萄藻生产的碳氢化合物可通过加氢裂化转化为柴油或喷气燃料,产率高达85%。使用藻类碳氢化合物可以大大减少使用煤炭和石油对环境的影响。与用于生物燃料生产的其它微藻相比,其涉及培养条件优化的工作相对较少。但与三酰甘油的生产不同,碳氢化合物的产量与布朗葡萄藻的生长有关,由于布朗葡萄藻的低生长速率,其生物量和碳氢化合物的生产力还有很大的提高空间。目前对布朗葡萄藻的研究进展突显了通过基因、光生物反应器及生物工程手段降低其生产成本的潜在可能,提高了其规模化培养及工业化生产的可行性。
04 展望
与展望本文从生物学特性、碳氢化合物合成途径及调控因子、多组学研究和规模化培养几方面叙述了布朗葡萄藻作为新型产油微藻生产碳氢化合物的潜力及应用价值。针对商业和工业应用领域的市场战略,应达到降低投资成本、提高利润的目的,使以藻类为原料的企业能够利用科学方法改进生物产品的生产。布朗葡萄藻能够产生环保的燃料及其他增值化学品,但仍需低成本高效益的提取技术和简单且可持续的规模化培养技术作为支持。对于布朗葡萄藻的基础研究能够帮助我们了解其生长性能,组学技术和系统生物学的发展使得人为操纵藻类的代谢途径成为可能,将藻类作为微生物细胞工厂应用的想法推到了前沿。微藻可以在实验室规模上生产少量目标产物,采用合适的生物光反应器、不同的实验条件及培养技术,可以提高微藻生物制品的产量和成本效益,并获得利润。但真正做到利用布朗葡萄藻工业化生产生物产品依旧面临着几方面的难题,如大规模生产生物质的低成本培养系统的建立、高生产力和产量的产物提取技术的优化、经济可行的下游处理以及残渣的再利用等等。





